Articles
- Page Path
- HOME > J Korean Soc Clin Toxicol > Volume 21(2); 2023 > Article
-
Original Article
SH-SY5Y 인간 신경모세포종 세포에서 MPTP 유발 세포 독성에 대한 거저리(Tenebrio molitor) 추출물의 보호효과 -
조인호1
 , 김유지2, 김선태1,3
, 김유지2, 김선태1,3
- Protective effects of mealworm (Tenebrio molitor) extract on N-methyl-4-phenyl-1,2,3,6-tetrahydropyridine (MPTP)–induced cellular toxicity in SH-SY5Y neuroblastoma cells
-
In Ho Jo, M.D.1, Yoo Ji Kim2, Seon Tae Kim, M.D.1,3
-
Journal of The Korean Society of Clinical Toxicology 2023;21(2):81-91.
DOI: https://doi.org/10.22537/jksct.2023.00021
Published online: December 29, 2023
- 422 Views
- 3 Download
1원광대학교병원 응급의학과
2원광대학교 의과대학 생리학교실
3원광대학교 의과대학 응급의학교실
1Department of Emergency Medicine, Wonkwang University Hospital, Iksan, Korea
2Department of Physiology, Wonkwang University School of Medicine, Iksan, Korea
3Department of Emergency Medicine, Wonkwang University School of Medicine, Iksan, Korea
- Corresponding author: Seon Tae Kim Department of Emergency Medicine, Wonkwang University Hospital, Wonkwang University School of Medicine, 895 Muwang-ro, Iksan 54538, Korea Tel: +82-63-859-1072 Fax: +82-63-859-2129 E-mail: kst5478@wku.ac.kr
Copyright © Korean Society of Clinical Toxicology
This is an Open Access article distributed under the terms of the Creative Commons Attribution Non-Commercial License (http://creativecommons.org/licenses/by-nc/4.0/) which permits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original work is properly cited.
Abstract
-
Purpose
- Edible insect extracts have been used as an alternative source for medicinal supplements due to their significant antioxidative and anti-inflammatory activity. Recent studies have reported that anti-microbial peptides from insects have neuroprotective effects on dopamine toxins. The purpose of this study was to investigate the protective functions of mealworm (Tenebrio molitor) extract (MWE) on N-methyl-4-phenyl-1,2,3,6-tetrahydropyridine (MPTP)–induced cellular toxicity in SH-SY5Y neuroblastoma cells.
-
Methods
- Cellular toxicity induced by the MPTP toxin and the impact of MWE on cell survival were analyzed using MTT assays. DAPI staining was performed to observe apoptotic phenomena caused by MPTP. Changes in caspase-3 activity and protein expression were observed using enzyme activity assays and western blot assays, respectively.
-
Results
- MWE exerted significant antioxidant activity, which was measured by both DPPH and ABTS radical assays, with a dose-dependent relationship. Furthermore, MWE resulted in cellular proliferation in SH-SY5Y cells in a dose-dependent manner. Furthermore, MWE pretreatment significantly inhibited MPTP-induced cytotoxicity, with a dose-dependent relationship. The morphological characteristics of apoptosis and increased reactive oxygen species induced by MPTP were also significantly reduced by MWE pretreatment.
-
Conclusion
- MWE treatment significantly attenuated MPTP-induced changes in the levels of proteins associated with apoptosis, such as caspase-3 and PARP. These findings suggest that MWE exerts neuroprotective effects on human neuroblastoma SH-SY5Y cells subject to MPTP-induced dopaminergic neurodegeneration.
- 파킨슨병(Parkinson’s disease, PD)은 퇴행성 뇌질환 중 2번째로 많으며, 노인인구가 증가하면서 발병률도 상승한다. 병인학적으로 중뇌 흑질 부위의 도파민성 신경세포의 사멸에 의해 신경전달물질 중의 하나인 도파민이 결핍되면서 운동완서, 진전, 강직 등의 운동장애가 발생된다1,2). 파킨슨병 환자에 대한 사후연구는 극히 어렵기 때문에 이 질병에 대한 여러 연구 모델이 개발되었는데, 신경독성을 이용하여 만든 모델에서 주로 수행되고 있으며 catecholamine성 신경만을 선택적으로 파괴하는 것으로 알려진 N-methyl-4-phenyl-1,2,3,6-tetrahydropyridine (MPTP)라는 물질을 이용한 파킨슨 질환 모델이 가장 많이 사용되고 있다3). In vitro 파킨슨병 모델로는 SH-SY5Y 신경세포주를 이용하여 MPTP에 의해 유도되는 세포사멸 현상이 널리 사용되며, 다양한 약물 및 약용식물의 항산화 물질 및 천연 폴리페놀 추출물은 파킨슨병 모델을 유발하는 MPTP에 의한 신경독성을 효율적으로 억제 또는 보호하는 효과가 있는 것으로 보고되었다4,5).
- 우리나라에서 메뚜기, 백강잠, 식용누에유충・번데기, 갈색거저리 유충, 쌍별귀뚜라미, 흰점박이꽃무지 유충, 장수풍뎅이 유충, 아메리카왕거저리 유충(탈지 분말), 수벌 번데기, 풀무치(‘21 한시적 인정) 등 총 10종이 식품원료로 사용 가능한 식용곤충으로 식약처로부터 인정받았다. 식용곤충은 저비용으로 사육할 수 있어 대체단백질 자원으로 활용이 가능하기 때문에 단백질 공급원으로써 곤충의 식용화와 관련된 산업이 전 세계적으로 급속도로 성장하고 있다. 식용곤충은 영양학적으로도 우수한 구성성분을 보유하고 있어 곤충을 활용한 기능성 소재를 개발하기 위한 연구가 활발히 이루어지고 있다6,7).
- 동양의학에서 식용곤충은 약용으로서 효용이 크기 때문에 질병치료 목적으로 오래 전부터 사용되어 왔으며, 우리나라에서는 동의보감 탕액편에 약 95종의 충부약재의 임상적 효능 및 주치가 서술되어 있다. 최근 들어 곤충 유래 한약재의 효능을 과학적으로 규명하고 기능성 소재로 개발하기 위한 연구가 활발히 이루어지고 있다. 특히 뇌신경계 질환을 타겟으로 한 연구가 다수 이루어져 관련 특허가 다수 등록되어 있으며, 이 중 선퇴(蟬退)의 항파킨슨 효과와 제조(蠐螬)의 뇌독성 억제효과 등이 보고되었다8,9).
- 갈색거저리(Tenebrio molitor)는 딱정벌레목 거저리과의 곤충으로, 갈색쌀거저리라고도 한다. 몸길이는 약 15 mm 정도이며, 몸 빛깔이 구룡충(구룡거저리)과 비슷해 어두운 갈색이며 광택이 난다. 갈색거저리의 유충은 밀웜(mealworm)이라 하고, 유충은 번데기가 되기까지 먹이와 온도에 따라 18–19번 탈피한다. 밀웜은 대두보다 필수 아미노산을 많이 함유하고 있고, 육류에 비해 불포화 지방산 함량이 높으며, 비타민 A와 철, 식이섬유 등이 비교적 풍부한 편으로 영양적인 장점이 많다10). 이와 관련하여, 갈색거저리가 단백질 함량이 매우 높은 고단백질 소재로 식품 원료로 가치가 높다고 평가되어 미래의 식량 자원으로 각광받고 있으며, 2016년 식용 곤충원료로 식품공전에 등록됨으로써 갈색거저리의 활용에 대한 관심이 높아지고 있지만6), 갈색거저리를 한방분야에서 약용으로 사용한 예는 드물다. 갈색거저리 추출물(mealworm extract, MWE)의 기능성 연구로는 항산화 및 항염증 효과, 골질환의 개선, 혈전증의 예방, 근육생성 촉진의 생리활성 작용들이 보고되었다10-14).
- 곤충유래 항균 펩타이드(antimicrobial peptide)는 바이러스 및 세균 감염으로 곤충을 보호하기 위하여 곤충에서 자체 합성되는 저분자 단백질체로, 항균, 항염증, 항산화 기능들을 가지고 있다15). 또한 곤충유래 항균 펩타이드들은 신경세포의 성장 및 사멸에 관여한다는 연구들이 보고되었으며, 특히 항균 펩타이드의 합성체가 펩타이드들이 SH-SY5Y 세포주의 분열을 촉진할 뿐만 아니라 MPTP에 의해 유도되는 in vitro 파킨슨 모델에서 신경세포의 사멸을 억제하다는 연구결과들이 보고되었다16,17). 그러나 MWE 자체가 in vitro 파킨슨 모델에서 신경세포의 사멸에 미치는 효과는 보고되지 않았다. 따라서 본 연구의 목적은 MPTP 신경독성에 의한 SH-SY5Y 세포주의 성장 및 사멸에 갈색거저리 수용성 추출물이 미치는 영향을 관찰하고자 하였다.
서론
- 1. 시약 및 항체
- Dulbecco’s Modified Eagle’s Medium (DMEM), fetal bovine serum (FBS), antibiotics-antimycotics 및 trypsin-ethylenediaminetetraacetic acid (EDTA) 등은 Thermo Fisher Scientific Inc. (Gibco, Grand Island, NY, USA)로부터 구입하였다. 2’,7’-dichlorodihydrofluorescein diacetate (DCFH-DA), 3-(4,5-dimethylthizol-2-yl)-2,5-diphenyl tetrazolium bromide (MTT), N-methyl4-phenyl-1,2,3,6-tetrahydropyridine (MPTP), sodium dodecyl sulfate (SDS), dimethyl sulfoxide (DMSO), 4′,6-diamidino-2-phenylindole (DAPI) 등은 Sigma-Aldrich Chemical (St. Louis, MO, USA)로부터 구입하였다. Nitrocellulose paper (Hybond-C, RPN303C)는 Amersham Biosciences Co. (Amersham, UK)에서 구입하였다. Anti-cleaved caspase-3와 anti-PARP (poly(ADP-ribose) polymerase) 항체 등은 Cell Signaling Technology Inc. (Beverly, MA, USA)로부터 구입하였으며, anti-β-actin 항체는 Santa Cruz Biotechnology (Santa Cruz, CA, USA)에서 구입하였다.
- 2. 세포배양
- 사람 신경모세포종 SH-SY5Y 세포주는 한국세포주은행(Korean Cell Line Bank, Seoul, Korea)에서 분양받았으며, 10% FBS와 1% antibiotics-antimycotics가 함유된 DMEM 배지를 사용하여 37℃와 5% CO2 조건에서 배양하였다. 일주일에 2번 간격으로 0.05% trypsin-EDTA를 사용하여 계대배양을 하였으며, 배양액을 교체한 후 log phase에 있는 세포를 사용하였다. 세포는 1×105 cells/mL로 seeding하여 24시간 전배양 후, 10% FBS가 포함된 DMEM 배지에서 MWE와 함께 MPTP을 처리하였다.
- 3. 항산화 효능 검사
- 1,1-diphenyl-2-picrylhydrazyl (DPPH) radical 소거능은 상추발효추출물을 아질산염(NO2-) 농도를 기준으로 10, 50, 100, 200 ppm 농도로 희석하여, 각각의 희석된 시료액 50 μL와 DPPH 용액 50 μL를 혼합하여 암실조건에서 30분간 방치시킨 후, SpectraMAX M2 multi-mode microplate reader (Molecular Devices, Sunnyvale, CA, USA)를 이용하여 517 nm에서 흡광도(optical density, OD)를 측정하였다. 시료를 첨가하지 않는 음성 대조군과 비교하여 유리라디칼 소거활성(radical scavenging activity)을 다음과 같이 계산하여 백분율(%)로 나타내었다.
- ABTS radical 소거능은 7.4 mM ABTS 용액에 2.6 mM K2S2O8를 혼합하여 암실조건에서 12시간 정도 반응시킨 후 734 nm에서 OD 값이 0.700±0.05가 되도록 ABTS 용액을 희석하여 사용하였다. ABTS 용액 180 μL와 각각의 희석된 시료액 20 μL를 혼합하여 암실조건에서 15분간 유지시킨 후, SpectraMAX M2 multi-mode microplate reader (Molecular Devices)를 이용하여 734 nm에서 OD를 측정하였다. 시료를 첨가하지 않는 음성 대조군과 비교하여 유리라디칼 소거활성을 다음과 같이 계산하여 백분율(%)로 나타내었다.
- 4. 갈색거저리 추출물 제조
- 본 연구에 사용한 갈색거저리는 스마트팜 시설에서 무균상태로의 곤충사육시설에서 사육하는 농업회사법인 흙농(Heuknong Co. Ltd., Namwon, Korea)에서 분양을 받았으며. 농촌진흥청에서 수립된 갈색거저리 사육 및 분말 제조 매뉴얼(Rural Development Administration 2015)에 근거하여 2일 이상 절식한 갈색거저리를 액체질소에 급속동결하고, –70℃ 초저온 냉동고(Nihon Freezer, Tokyo, Japan)에서 24시간 이상 냉동시켰다. 40 g 동결 갈색거저리를 0.1% acetic acid 함유 50 mM phosphate buffer (pH 7.4) 용액 100 mL (weight/volume)에 넣고 고속 분쇄기로 3분 동안 분쇄하였다. 갈색거저리 분쇄액을 다시 초음파 분쇄기에서 3분간 다시 추가적으로 분쇄하였다. 갈색거저리 분쇄액을 4℃에서 1일간 방치한 후 12,000 rpm으로 원심분리하여 상층액만 회수하여 실험에 사용하였다. 수용성 상층액(MWE)을 다시 PBS 용액에 3배 희석하여 세포실험에 사용하였다.
- 5. 세포 증식 및 생존율 측정
- SH-SY5Y 세포의 증식률은 세포를 24-wells 플레이트에 1×105 cells/mL씩 분주하여 24시간 전 배양하고 MWE을 5, 10, 25, 50 μL 용량으로 각각 처리하고 5일간 배양하였다. 배양 5일째 MTT (0.5 mg/mL) 용액을 첨가하고 37℃에서 4시간 배양하였다. 상등액을 주의 깊게 제거하고 살아있는 세포에 의해 생성된 보라색 불용성 formazan은 DMSO로 용해하였다. Formazan의 양은 SpectraMAX M2 multi-mode microplate reader (Molecular Devices)를 이용하여 570 nm에서 OD를 측정하였다. 세포생존율(cell viability)은 다음과 같이 계산하여 백분율(%)로 표시하였다.
- MPTP에 의한 SH-SY5Y 세포의 생존율 변화는 전 배양된 SH-SY5Y 세포에 MPTP를 1 mM과 2 mM 농도로 12시간 처치한 다음 MTT assay를 실시하였으며, MPTP에 의한 신경독성에 MWE의 효과는 전 배양된 세포에 5, 10, 25, 50 μL 용량으로 1시간 전처리한 다음 MPTP를 1 mM 또는 2 mM 농도로 12시간 처치한 다음 MTT assay를 실시하였다.
- 6. DAPI 염색
- 세포는 DAPI-메탄올(1 μg/mL) 용액으로 세척하고, DAPI-메탄올 용액으로 37℃에서 15분간 염색하였으며, 염색된 세포는 메탄올로 세척하고 Olympus IX71 형광현미경(Olympus, Tokyo, Japan)을 이용하여 세포 핵의 형태 변화를 관찰하였다.
- 7. 세포 내 활성산소종 측정
- MPTP에 의한 세포 내 활성산소종(reactive oxygen species, ROS)의 생성을 측정하기 위하여 형광 probe DCFH-DA를 이용하였다. 비형광 물질인 DCFH-DA는 세포 내 ROS와 반응하여 형광을 발하는 2',7'-dichlorofluorescein (DCF)로 변환되어 녹색의 형광을 나타낸다. 세포 내 ROS의 양을 측정하기 위하여 SH-SY5Y 세포를 배양하고 2 mM MPTP 및 5와 25 μL MWE를 6시간 처리한 후, 20 μM DCFH-DA를 첨가하여 37℃에서 30분간 배양 후 PBS (pH 7.4)로 세척하고 Olympus IX71 형광현미경(Olympus)으로 세포 내 ROS의 생성 정도를 관찰하였다. 이후 DMSO 용액으로 세포를 용해하여 세포 내 DCF 형광량을 SpectraMAX M2 multi-mode microplate reader (Molecular Devices)를 이용하여 excitation 파장 485 nm와 emission 파장 524 nm에서 측정하였다.
- 8. Caspase-3 활성 측정
- Caspase-3 활성측정은 Colorimetric Caspase-3 Assay Kit (Abacam, Cambridge, MA, USA)를 이용하여 측정하였다. 제조사의 방법에 따라서 세포용해물을 추출하였고, Bradford 검사법을 이용하여 단백질농도를 측정하였다. 50 μL의 세포용해물(총 50 μg의 단백질)과 50 μL의 2× reaction buffer 및 5 μL의 4 mM DEVD-pNA substrate를 혼합하여 37℃에서 1시간 배양하였다. Enzyme-catalyzation 되어 방출된 p-nitroaniline를 SpectraMAX M2 multi-mode microplate reader (Molecular Devices)를 이용하여 405 nm에서 측정하였으며, caspase-3 활성은 다음과 같이 계산하여 백분율(%)로 나타내었다.
- 9. 면역형광염색
- 면역형광염색을 위하여 SH-SY5Y 세포는 cover slip (Marienfeld, Lauda-Königshofen, Germany)을 넣은 24-well plate (Falcon; BD Biosciences, Franklin Lakes, NJ, USA)에서 배양한 상태에서 MWE 10–25 μL 용량을 처치하고 5일간 방치하였다. 배양액을 제거하고 PBS (pH 7.4)로 3회 세척을 하고, PBS에 희석시킨 4% paraformaldehyde (pH 7.2) 액으로 상온에서 10분간 고정을 하였다. PBS로 고정액을 세척하여 낸 후, PBS로 희석한 0.1% Triton X-100 액에 anti-Ki67 항체(1:500, rabbit-polyclonal; Abcam, Boston, USA Iowa)를 1:300의 농도로 희석하여 SH-SY5Y 세포와 같이 24시간 동안 상온에서 반응시켰다. 반응을 마친 SH-SY5Y 세포는 PBS로 3회 세척하고, 다시 PBS에 1:500의 농도로 희석한 Alexa Fluor 488 anti-rabbit immunoglobulin G (Molecular Probes, Eugene, OR, USA)와 함께 1시간 동안 상온에서 반응을 시켰다. 핵 염색을 위하여 DAPI (1:5000; Molecular Probes)로 15분간 상온에서 염색을 하였다. 염색된 세포는 Olympus IX71 형광현미경(Olympus)을 이용하여 관찰하였다.
- 10. Immunoblot 분석
- 세포는 ice-cold PBS (pH 7.4)로 세척하고 RIPA lysis buffer로 얼음 위에서 30분간 용해한 후, 세포용해물들은 4℃, 14,000 rpm에서 15분간 원심분리하였다. 추출한 단백질의 농도는 Bradford 단백질 정량법을 이용하여 측정하였고 동량의 단백질을 SDS–polyacrylamide gel electrophoresis로 전기영동하였다. 전기영동한 gel에서 nitrocellulose membrane에 단백질을 전이시켰고, 5% skim milk가 함유된 Tris-buffered saline + Tween-20 (20 mM Tris-HCl, 150 mM NaCl, pH 7.6, 0.05% Tween-20; TBS-T)로 비특이적 결합을 차단시켰다. 그 후 단백질의 발현을 분석하기 위해, 1차 항체(1:1,000)를 반응시켰으며, peroxidase가 붙어 있는 2차 항체(1:2,000)로 1시간 처리한 다음 TBS-T로 세척을 한 후 ECL kit로 1분간 반응시켜 FUSION Solo S (Vilber Lourmat, Marne-la-Vallée cedex, France)을 이용하여 특정 면역반응 단백질 밴드를 시각화하였다. 단백질 밴드의 강도는 Quantity ONE ver. 4.6.6 software (Bio-Rad, Hercules, CA USA)를 이용하여 정량화하였다.
- 11. 통계 처리
- 표시된 결과는 3회 이상의 독립적인 실험결과를 사용하였으며 GraphPad Prism software ver. 6.0 (GraphPad Software, San Diego, CA, USA) 프로그램을 통하여 통계 처리하여 mean±standard deviation으로 기록하였다. 실험군 간의 유의성 검정은 Student t-test를 통해 분석하였으며, p<0.05의 경우에서 통계적 유의성이 있는 것으로 간주하였다.
대상과 방법
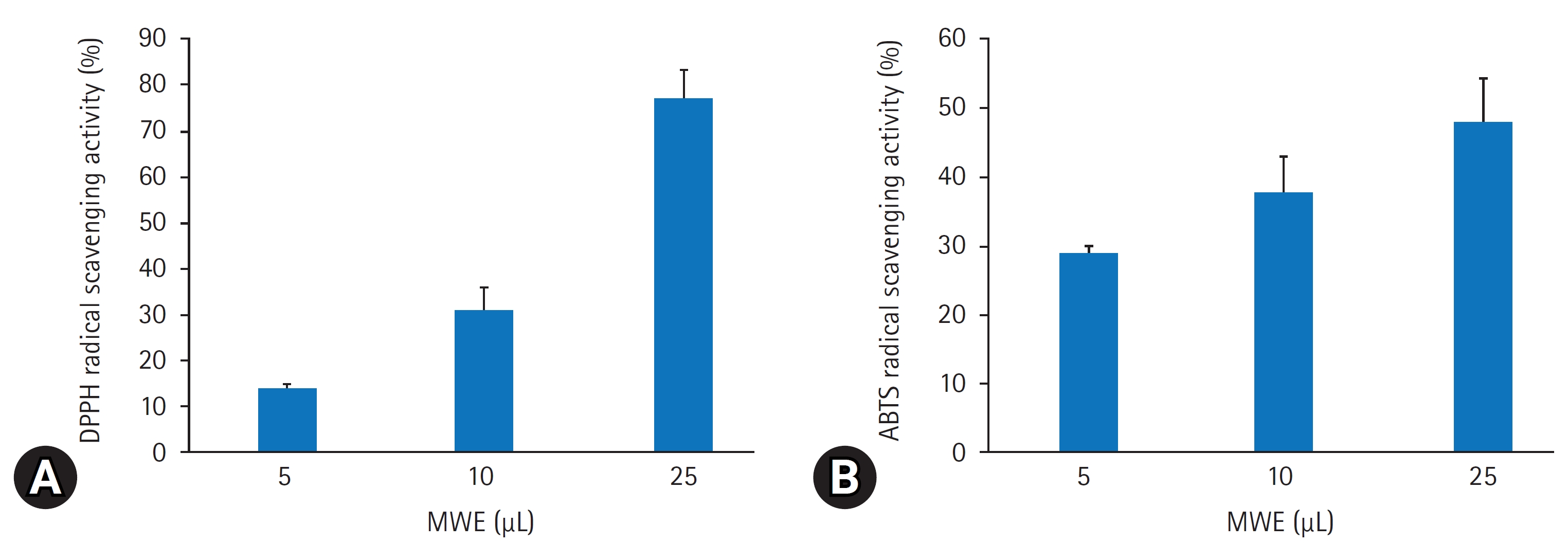
- 1. 갈색거저리 추출물의 항산화 효과
- DPPH 라디칼 소거법은 항산화 물질의 전자공여능으로 인해 방향족 화합물 및 방향족 아민류에 의해 환원되어 자색이 탈색되는 정도를 지표로 하여 항산화능을 측정하는 방법이다. 본 실험에서는 MWE의 추출물을 용량별(5–25 μL)로 DPPH에 대한 라디칼 소거활성을 측정한 결과, 용량 의존적으로 증가하는 경향을 보였으며 25 μL의 농도에서 77.3%6.3%의 소거능을 보였다(Fig. 1A). 한편, ABTS법에 MWE의 ABTS 라디칼 소거능력은 DPPH 소거능과 비슷하게 용량 의존적으로 증가하여 25 μL 용량에서 47.9%1.1%의 소거능을 보였다(Fig. 1B).
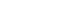
- 2. 갈색거저리 추출물의 SH-SY5Y 세포증식 효과
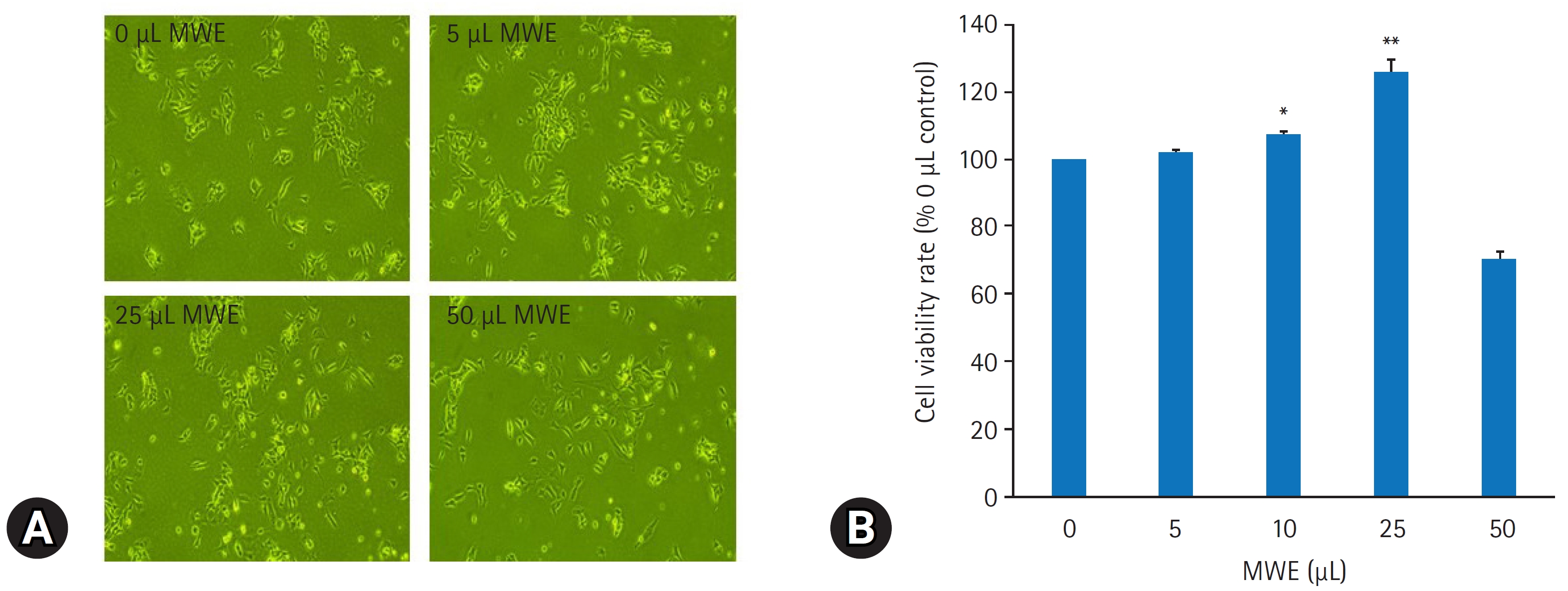
- MWE의 SH-SY5Y 세포의 증식에 미치는 효과를 평가하기 위해 MWE을 5–50 μL 용량으로 처리하고 6일 후 MTT 검사를 통해 세포생존율을 측정하였다. MWE은 5–25 μL까지 용량 의존적으로 세포생존율이 증가하였으며, 특히 25 μL 투여 시 음성 대조군(0 μL)과 비교하여 125.8%3.3% 증가하였다(p<0.01). 그러나 50 μL 용량의 MWE 처치한 경우는 세포생존율이 감소되었다(Fig. 2B). MWE의 세포증식 효과를 재확인하기 위해 MWE를 72시간 처리한 후, 면역형광 염색을 통해 세포분열의 지표단백질인 Ki67 단백질 발현을 관찰하였다. 영상분석상 200배 현미경 시야에서 MWE을 처치하지 않은 대조 세포들에 Ki67 양성 세포 수는 20.54.7이었으며 25 μL MWE을 처치하지 하였을 때 Ki67 면역 양성 세포 수가 33.25.2로 통계적으로 유의하게 증가하였다(p<0.05) (Fig. 3).
- 3. MPTP로 유도된 SH-SY5Y 세포의 신경독성에 대한 갈색거저리 추출물의 효과
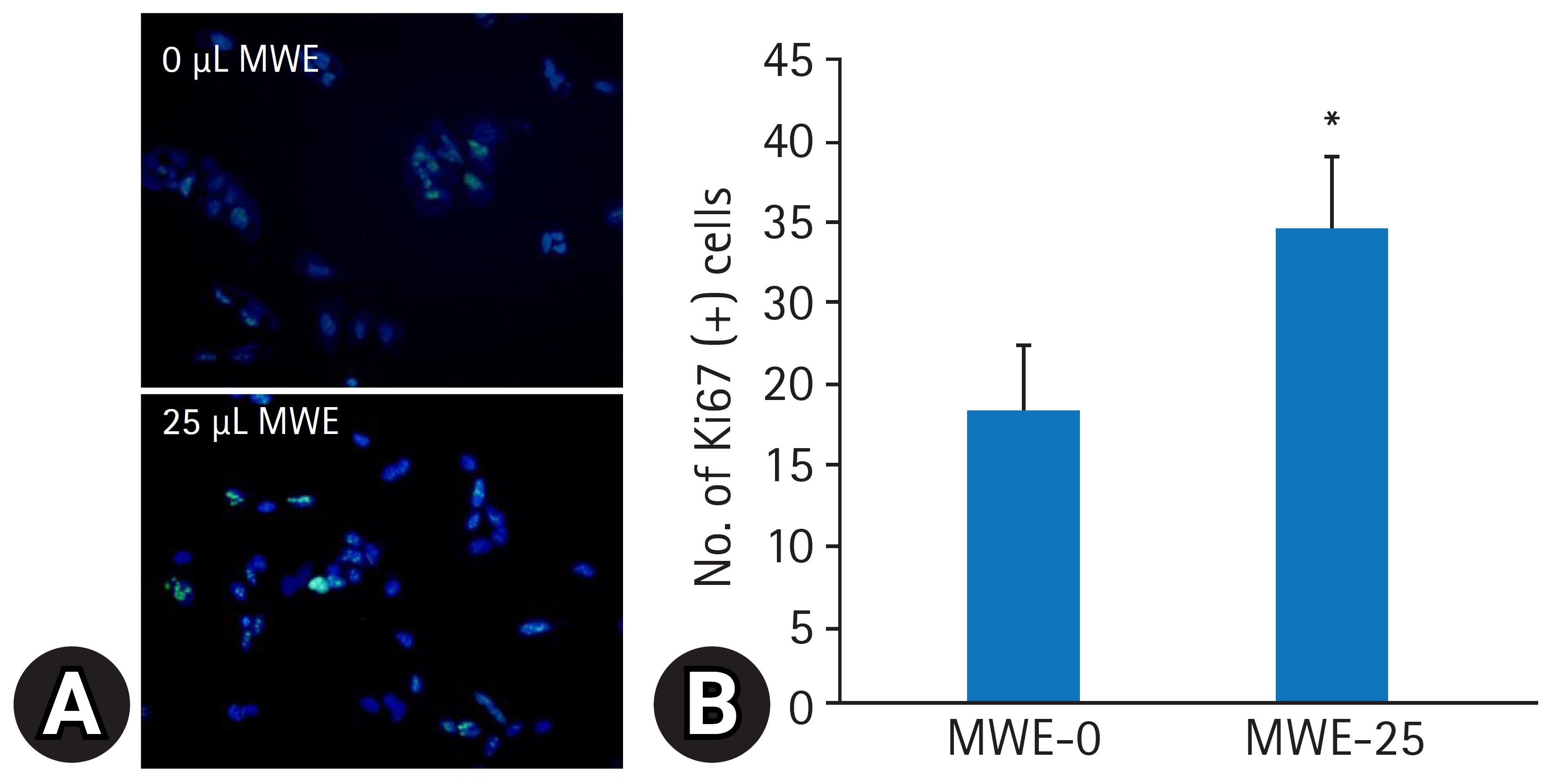
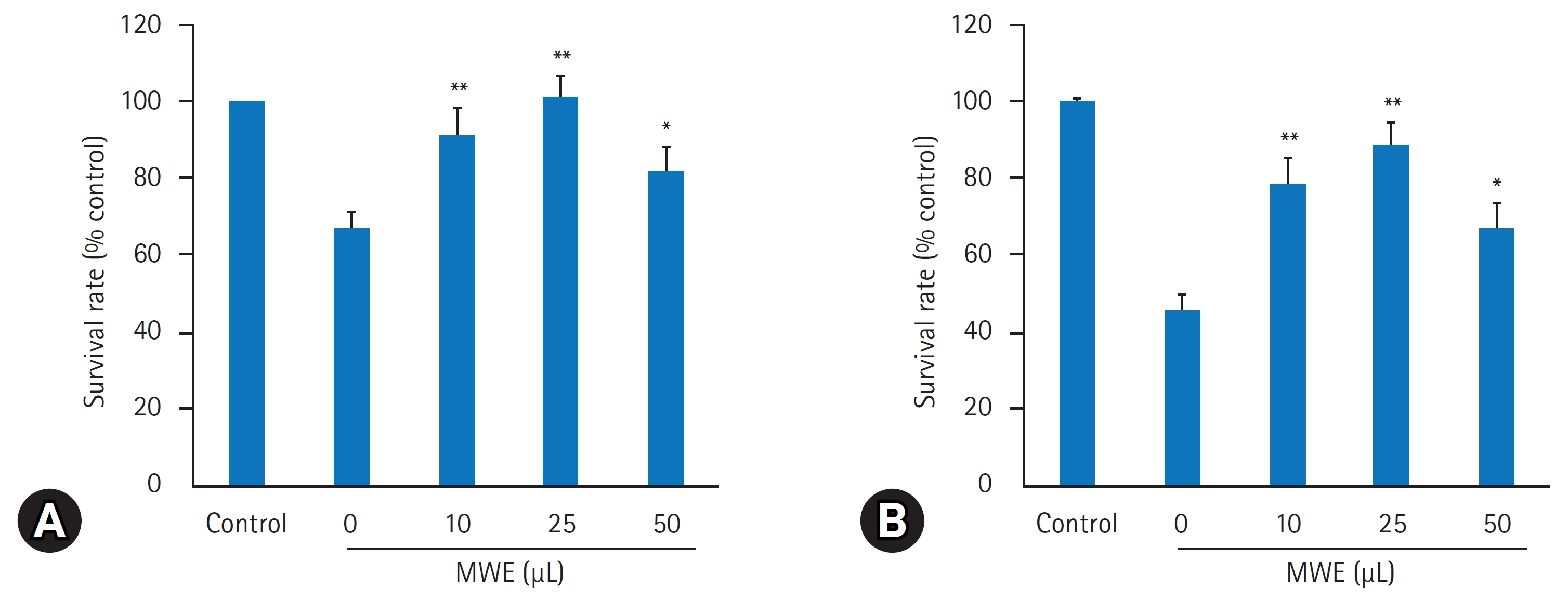
- MPTP의 산화성 손상으로 유발되는 신경독성에 의한 SH-SY5Y 세포의 생존율을 평가하기 위해 1 mM과 2 mM 농도로 MPTP를 12시간 처리한 후, 현미경적 관찰 및 MTT 검사로 세포생존율을 조사하였다. MTT 검사에서 MPTP 농도가 증가할수록 세포생존율이 농도의존적으로 유의하게 감소되어 1 mM MPTP 처치 시 MPTP을 처치하지 않은 음성 대조군의 생존율(100%) 대비 67.25.3, 2 mM MPTP 처치 시 45.2%4.5%의 세포생존율을 보였다(Figs. 4, 5). MPTP의 신경독성에 의한 SH-SY5Y 세포의 생존율 감소에 대한 MWE의 효과를 조사하기 위해 SH-SY5Y 세포에 10–50 μL 용량으로 MWE을 1시간 전처리하고 1 mM 또는 2 mM 농도로 MPTP를 12시간 처리한 후 세포생존율을 측정하였다. MPTP만 처치한 실험군보다 10 μL과 25 μL MWE와 MPTP를 동시에 처치한 실험군에서 MWE는 1 mM 및 2 mM 농도의 MPTP 처치한 양쪽 모두에서 세포생존율이 용량 의존적으로 유의하게 증가하였다(p<0.01). 특히 2 mM MTPT 처치한 세포에서 10 μL과 25 μL MWE는 세포생존율을 각각 78.2%7.1%와 88.2%5.5% 통계적으로 유의하게 증가시켰다(p<0.01). 한편, 2–5 μL 용량의 MWE는 MPTP에 의해 감소된 세포생존율을 유의하게 증가시키지 못하였으며, 50 μL 용량에서는 세포생존율 감소를 악화시켰다(p<0.05) (Figs. 4, 5).
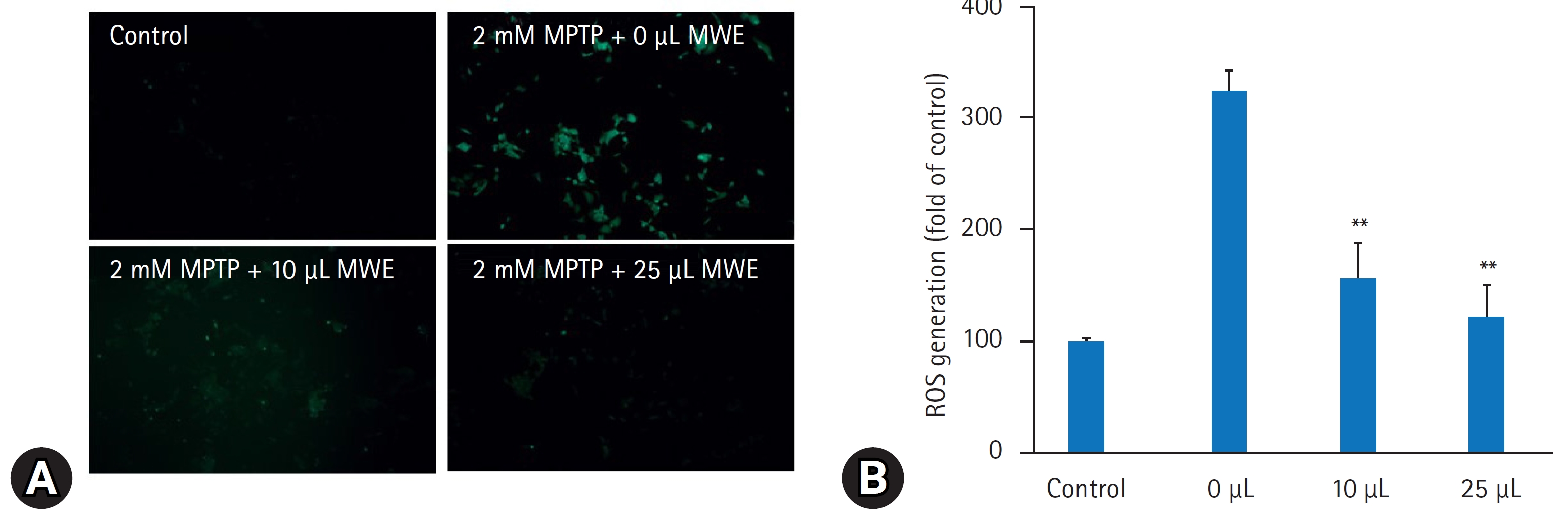
- 4. MPTP에 의한 SH-SY5Y 세포 내 ROS 생성에 대한 갈색거저리 추출물의 효과
- MPTP는 세포 내 미토콘드리아 기능 장애를 초래하여 ROS를 발생하여 산화성 스트레스 손상을 초래하는 것으로 알려져 있다. MPTP에 의한 세포 내 ROS 생성을 검출하기 2 mM MPTP 처치 6시간에 DCFH-DA를 처치하고 2시간 후에 형광현미경으로 관찰하였다. MPTP을 처치하지 않은 대조실험군의 세포에서는 ROS에 의한 DCF 형광이 잘 관찰되지 않았으나 MPTP만 처치한 세포에서 ROS에 의한 DCF의 형광량이 유의하게 증가하였으며 형광 spectrophotometer로 실험군 간에 DCF의 형광량을 정량하였을 때 음성 대조군의 DCF 형광량과 비교하여 MPTP만 처치한 경우 325.1%20.2% 증가하였다(p<0.01). 그러나 10 μL과 25 μL MWE을 전처리한 경우 MPTP에 의해 증가된 DCF의 형광량이 유의하게 감소하였으며 2.5% (volume/volume) MWE 처치 시 대조군의 발현량과 비교하여 120.5%32.8%로 MPTP만 처치한 경우와 비교하여 통계적 유의하게 감소하였다(p<0.01) (Fig. 6).
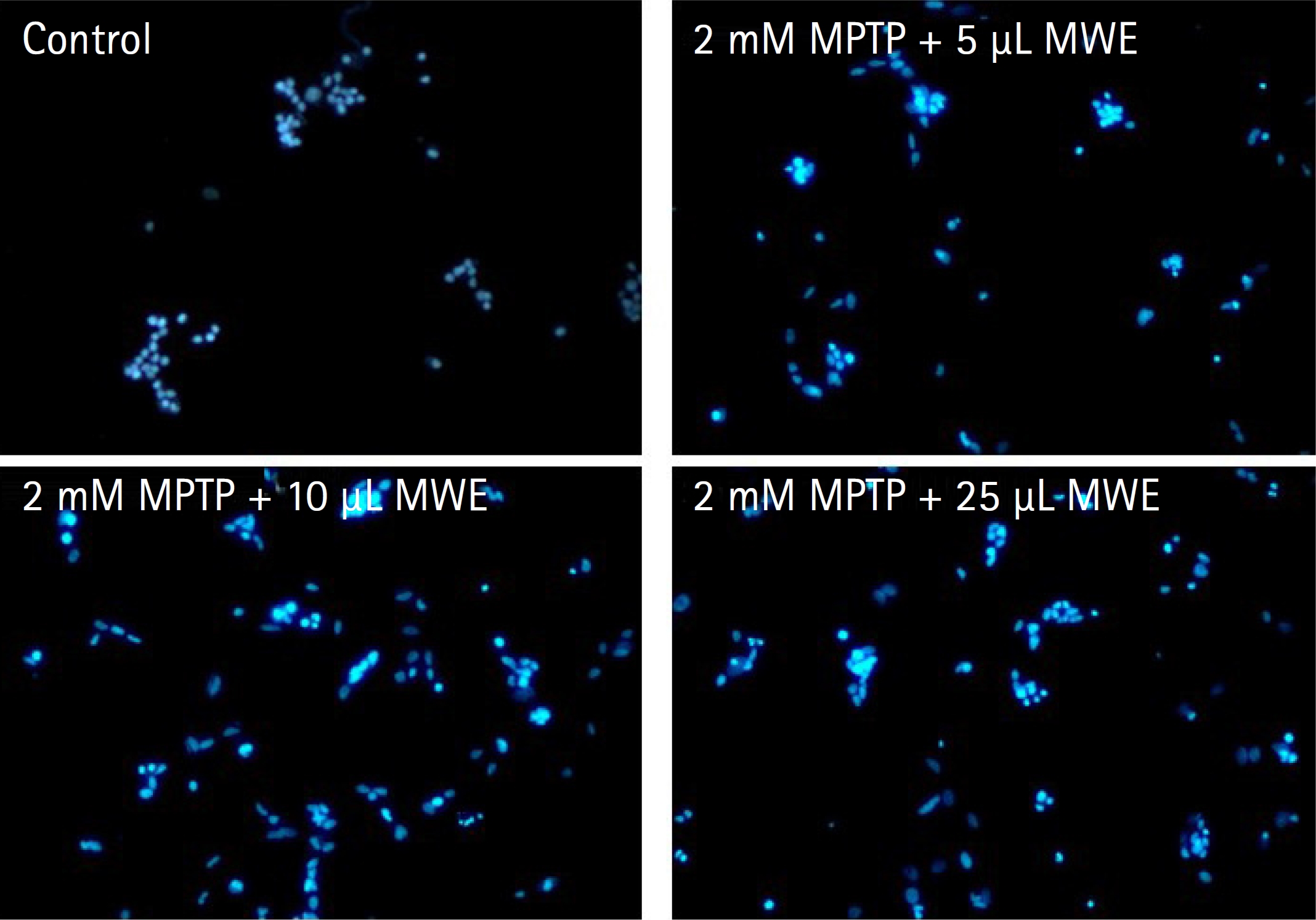
- 5. MPTP 신경독성에 의한 SH-SY5Y 세포자멸사에 대한 갈색거저리 추출물의 효과
- MPTP로 유도된 SH-SY5Y 세포자멸사에 대한 MWE의 효과를 조사하기 위해 DAPI 염색을 통해 세포 핵의 형태변화를 관찰한 결과 Fig. 7에서 나타난 바와 같다. MPTP를 처치하지 않은 음성 대조군의 세포 핵에서는 DAPI 염색이 연하고 균일하게 관찰되었지만, 2 mM MPTP를 처치하고 12시간 경과된 세포에서는 핵들이 응축(condense)되면서 DAPI의 투과성 증가에 따른 빛의 강도증가 현상과 핵들의 분절에 세포자멸소포(apoptotic body)의 형성이 저명하게 관찰되었다. 그러나 MWE를 처치한 경우 농도의존적으로 이러한 세포자멸사(apoptosis) 유발에 의한 핵들의 형태학적 변화가 감소하였으며, 특히 5–25 μL 농도에서 더욱더 감소하였다(Fig. 7).
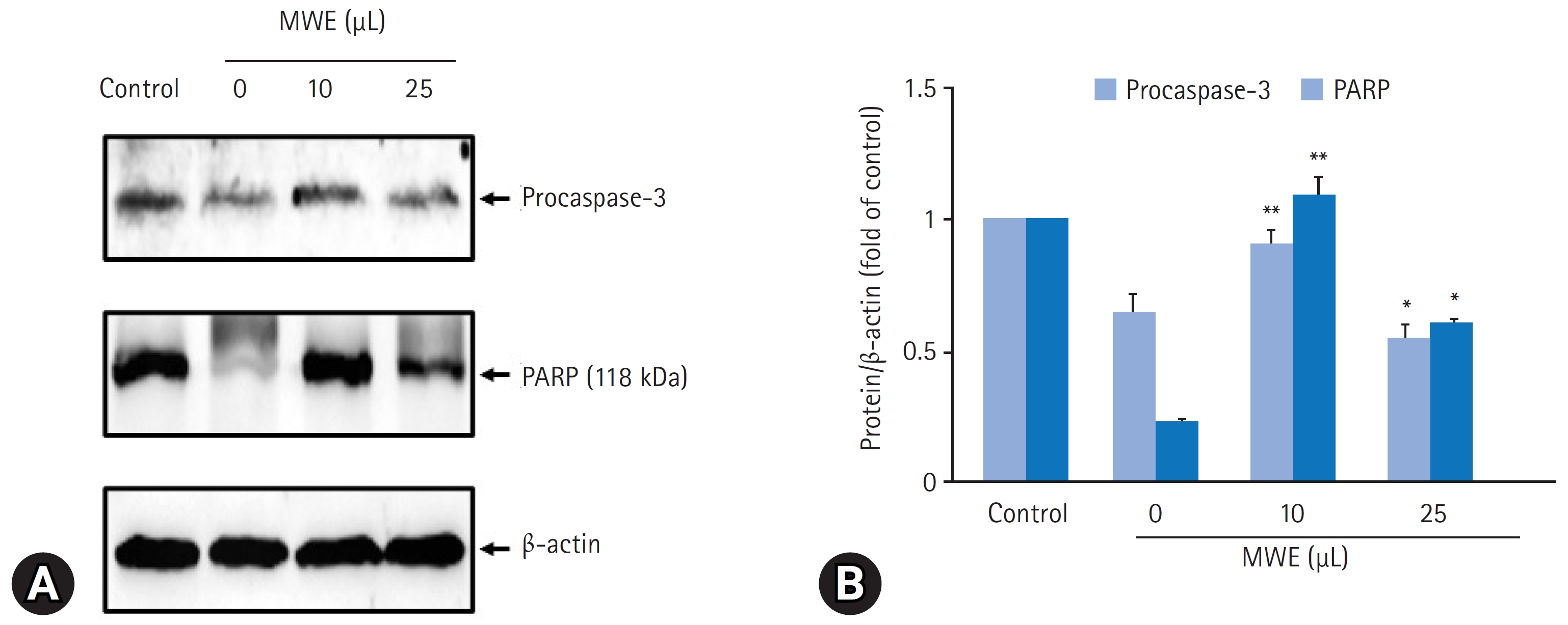
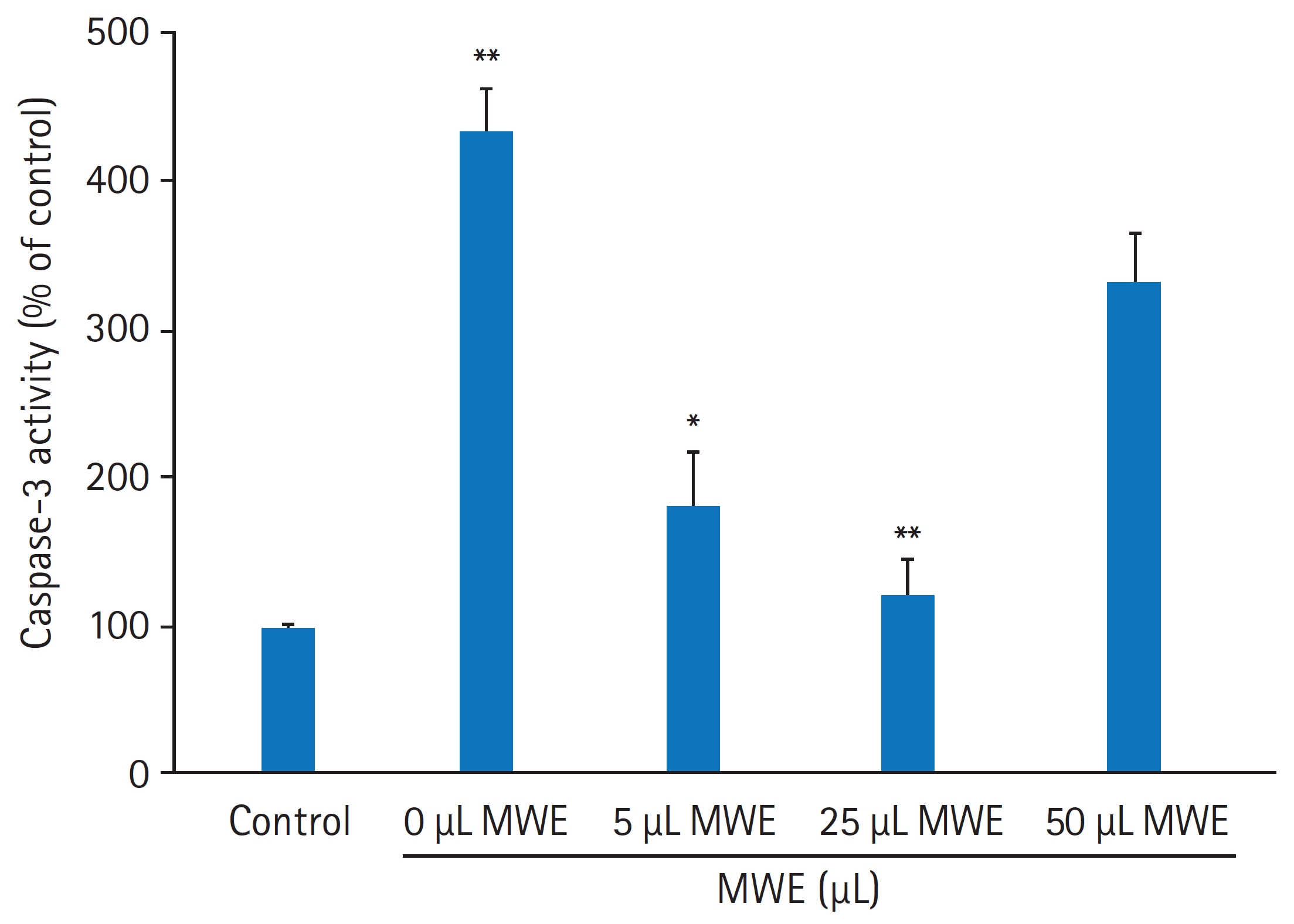
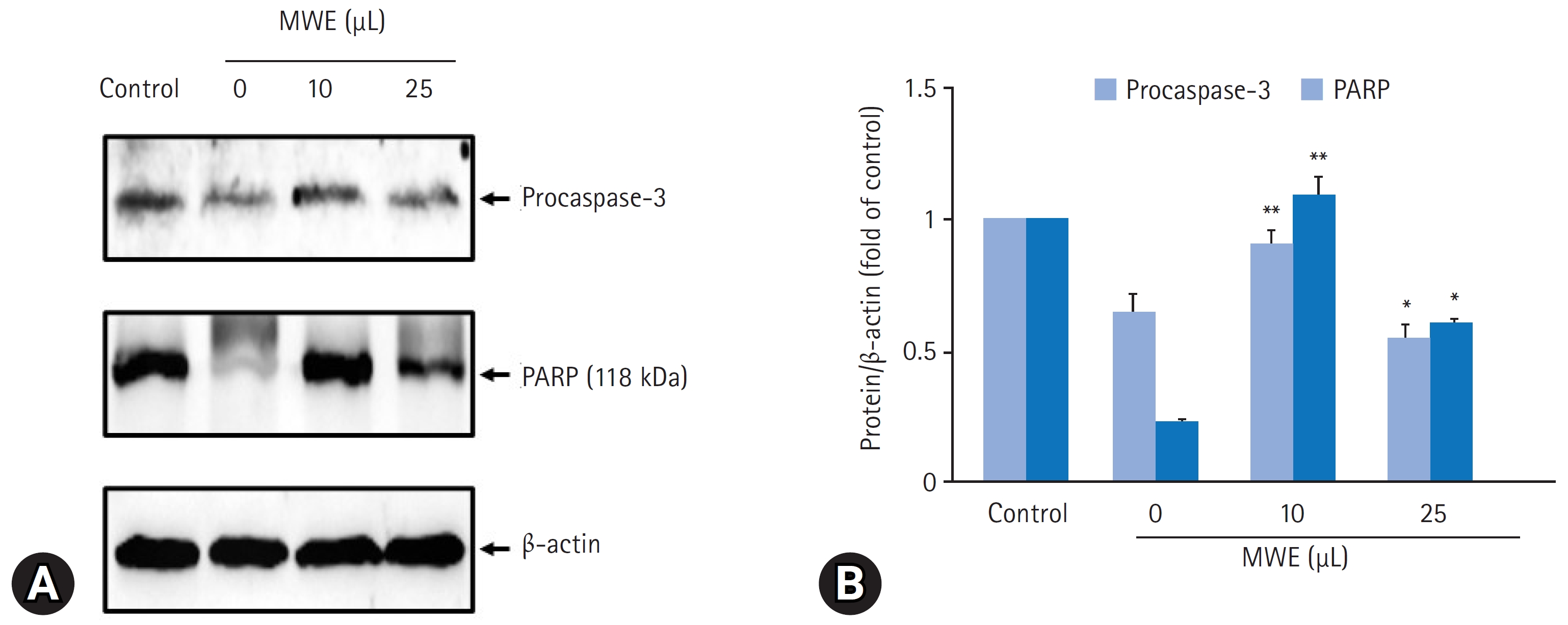
- MPTP로 유도된 SH-SY5Y 세포자멸사에 대한 MWE의 억제효과를 재검증하기 위해 세포자멸사의 지표단백질인 caspase-3 활성 및 불활성형 procaspase-3 발현과 caspase-3 활성에 의한 PARP (118 kDa) 발현 변화를 조사하였다. MPTP을 처치하지 않은 음성 대조군의 세포와 비교하여 2 mM MPTP를 처치하고 24시간 경과된 세포에서 caspase-3의 활성은 432.7%25.4%로 통계적으로 유의하게 증가하였다(p<0.01). 그러나 10 μL와 25 μL MWE를 전처치하고 2 mM MPTP 처치하였을 때 caspase-3의 활성은 통계적으로 유의하게 감소하여 25 μL MWE를 전처치한 실험군에서 120.3%25.2%였으나, 50 μL MWE를 전처치하고 2 mM MPTP를 처치하였을 때 caspase-3의 활성은 음성 대조군의 330.1%33.3%로 증가되어 MPTP만 처치한 군과 통계적 차이를 보이지 않았다(Fig. 8). 2 mM MPTP만 처치한 세포에서 procaspase-3 및 PARP 발현은 음성 대조군(control)의 세포와 비교하여 유의하게 감소한 반면, 10 μL 또는 25 μL MWE를 전처치하였을 경우 MPTP에 의해 감소된 procasaspe-3 및 PARP 발현이 유의하게 증가되었다(Fig. 9).
결과
- 곤충은 동양의학에서 약용으로써 질병치료 목적으로 사용되어 왔으며 약용곤충은 약용식물과의 혼합을 통하여 이용되었고, 다양한 가공방법으로 가공하여 사용하였다. 중국에서는 약 2천 년부터 사용하였으며, 곤충의 약용으로써 이용은 ‘주기(周記)’, ‘산해경(山海經)’, ‘신농본초경(神農本草經)’, ‘명의별녹(名醫別錄)’, ‘본초(本草經集主)’, ‘본초강목(本草綱目)’ 등에 의해 확인되고 있다. 일본의 ‘약용곤충목록(藥用昆蟲目錄)’, ‘곤충 본초’ 등 또한 약용으로 이용할 수 있는 곤충을 설명하고 있다. 우리나라에서는 ‘동의보감’에 의해 약 95종의 곤충이 약용으로 쓰일 수 있다고 서술되어 있으며, 일부 문헌에서 발췌된 기록을 통해 역사적 고증을 거쳐 식용 및 약용곤충의 선정에 활용하고 있다6,8).
- 갈색거저리 유충은 고단백 미래 식량자원으로 각광받고 있으며, 건강기능식품과 의약품으로도 활용가치가 높은 것으로 알려져 있다. MWE의 기능성 연구에서 세포 성장, 항염증, 항산화 효과에 지방간 개선, 골질환의 개선, 혈전증의 예방, 근육생성 촉진 등에 관여하는 것으로 알려져 있다6,14,18,19).
- 본 실험에서 갈색거저리 수용성 추출물(MWE)에 항산화, 항염증 등의 효과를 나타내는 폴리페놀(polyphenol) 화합물은 100 mg gallic acid equivalent (GAE)/100 mL의 농도를 보였다. Mattia 등(2019)20)도 갈색거저리 수용성 추출물에서 총 폴리페놀 함량은 300 mg GAE/100으로 비교적 곤충들 중에 많은 것으로 보고하였다. 또한 본 실험에서 MWE는 DPPH 및 ABTS에 대한 라디칼 소거활성을 측정한 결과, 농도의존적으로 증가하는 경향을 보여 25 μL 용량에서 77.3%의 소거능을 보였다. 갈색거저리 유충 단백 가수분해물도 DPPH 라디칼 소거활성에 50% 라디칼 소거율을 보였다21,22). 갈색거저리의 단백성분을 제거한 추출물도 DPPH에 대해 80% 라디칼 소거활성을 보였다23). 따라서 본 연구결과와 선행연구결과들을 종합하였을 때 갈색거저리의 단백과 비단백 성분의 폴리페놀 성분들이 항산화 작용을 가지고 있음을 보여준다.
- 본 연구에서 MWE는 용량 의존적으로 MTT 검사에서 OD 값이 증가하여 세포성장이 촉진되었다. 더불어 세포분열의 단백질 표지자인 Ki67 단백을 면역형광 염색하였을 때 MWE를 처치하지 않은 세포들에 비하여 25 μL MWE를 처치한 세포들에 Ki67 면역 양성 세포 수가 통계적으로 유의하게 증가하였다(p<0.05). MWE는 모모세포(human keratinocyte), 모유두세포(human dermal papilla cell), 섬유아세포(fibroblast) 및 지방세포에서 세포생존율과 세포증식 효과, 탈모 관련 사이토카인의 조절능력이 우수하다11,12).
- 곤충은 상처와 감염에 대응하여 다양한 혈액 및 임파액에서 세크로핀(cecropin) 및 디펜신(defensin) 계열의 항세균 펩타이드(antibacterial peptide)를 생성하며 세균 및 진균(fungus)에 대한 강력한 항균, 항진균 작용을 통해 생명을 유지한다24). 갈색거저리에는 아타신(attacin), 디펜신(defencin), 테네신(tenecin) 및 콜레옵테리신(coleoptericin)와 같은 항균단백질이 있으며 유산균 세균 추출물을 곤충의 배면에 주사기로 주입하여 “면역화(immunization)”를 유도하면 4–15시간 내에 항균펩타이드의 messenger RNA 및 단백 발현이 현저하게 증가된다25).
- 한편, 이러한 항균펩타이드들이 신경세포의 생존 및 사멸에 미치는 영향에 대한 여러 선행연구결과들이 보고된 바 있다. 즉 면역화된 쇠똥구리 곤충에서 유래한 항균펩타이드인 코프리신(Coprisin) 펩타이드의 핵심 구조 펩타이드인 CopA3 펩타이드는 사람 신경모세포종(neuroblastoma)에서 유래한 SH-SY5Y 도파민 세포와 대장세포종의 분열을 용량 의존적으로 촉진하였으며, SH-SY5Y 세포 및 신경줄기세포에서 시간 의존적으로 30% 이상 세포증식을 촉진하였다. 이때 세포 내 세포분열 억제 단백인 p27Kip1을 분쇄하여 세포분열을 촉진한다16). 지렁이에서 유래한 항균펩타이드인 lumbricusin (NH2-RNRRWCIDQQA)도 SH-SY5Y 도파민세포의 분열을 촉진하여 세포성장을 유도하였다17). 따라서 본 연구에서 사용한 갈색거저리에 함유된 항균펩타이드들이 신경세포의 분열 및 성장에 관여할 수 있음을 가정할 수 있으며, 본 연구에서 사용한 MWE 내 항균펩타이드들의 기전에 대한 추가 연구가 필요하다.
- MPTP에 의한 도파민 신경세포의 손상기전에 대한 다양한 연구가 진행되어 왔다. 신경세포의 미토콘드리 내 NADH와 관련된 산화성반응과 NADH dehydrogenase 활성도가 MPTP에 의해 억제되어 ATP 생성이 저해된다. 또한 MTPT의 대사물은 반응성이 높은 전자친화 대사물로 생체의 각종 중요한 거대분자와 작용하여 거대분자의 기능을 억제하며, 생체 내에 반응성 산소종의 생성을 초래하여 세포 내 지질, DNA 등을 손상시켜 세포사멸을 초래한다3).
- 본 연구에서 MPTP 처치 후 24시간 경과된 세포들에서 핵들이 응축되면서 DAPI의 투과성 증가에 따른 빛의 강도증가 현상과 핵들의 분절에 세포자멸소포(apoptotic body) 형성이 저명하였다. 그러나 MWE을 처치한 경우 용량 의존적으로 이러한 세포사멸 현상과 관련된 세포 핵의 형태학적 변화가 감소하였으며, 특히 10–25 μL 용량에서 유의하게 감소하였다. 더욱이 세포의 자멸사 과정에서 가장 공통적으로 관여하는 caspase-3 활성은 MPTP의 처치에 의하여 증가되었으나, 10 μL 및 25 μL MWE을 전처치에 의하여 유의하게 감소하였다. 그리고 MPTP만 처치한 세포에서는 ROS에 의한 DCF-DA의 형광량이 유의하게 증가하였으나 10–25 μL MWE를 전처리한 경우에는 MPTP에 의해 증가된 DCF-DA의 형광량이 유의하게 감소하였다. 또한 본 연구결과에서 MWE가 DPPH 라디칼 소거율을 증가시킴으로써 항산화 능력이 검증되었다.
- 정상적인 세포들은 호흡과정 동안 다양한 ROS (O2-, H2O2, OH- 등)을 내지만 이는 세포 자체가 갖고 있는 항산화시스템(glutathione, superoxide dismutase, catalase 등)에 의해 조절된다. 그러나 다양한 자극에 의해 과량의 ROS가 발현되었을 경우에는 세포 자체에 해로운 신호전달의 시발점이 되어 결국 세포사멸로 이어지게 되고, 분자생물학적 변화와 세포손상을 일으킨다26). 여러 연구에서 항산화 성분을 함유하고 있는 천연 폴리페놀 추출물들이 파킨슨병 모델을 유발하는 MPTP에 의한 세포독성을 효율적으로 억제 또는 보호한다는 결과가 보고되었다26,27). 따라서 MWE에 포함되어 있는 항산화 물질이 MPTP에 의한 신경세포 내 반응성 산소종의 생성을 억제하여 산화성 손상을 감소시킬 수 있음을 의미한다.
- 단백질을 포함하지 않는 MWE가 미세아교세포종인 BV-2 세포에서 lipopolysaccharide를 처리하여 증가되는 염증성 단백인 cyclooxygenase-2 (COX-2) 와 inducible nitric oxide synthase 단백 발현을 유의하게 억제하였다28). MPTP에 의한 in vitro 및 in vivo 파킨슨병 모델에서 선택적 cyclooxygenase (COX)-2 억제제는 행동학적 장애를 호전시켰으며 미토콘드리아 기능장애, caspase-3 및 nuclear factor kappa B 단백의 발현 감소를 보여 염증성 반응이 MPTP에 의한 도파민 세포사멸에 중요함을 밝혔다29,30). 따라서 선행연구결과들을 종합하였을 때 본 연구에서 사용한 MWE가 염증반응을 억제할 것으로 추정되며, 이에 대한 추가 연구가 필요할 것으로 생각된다.
고찰
- 본 연구는 SH-SY5Y 신경모세포종을 대상으로 신경세포 성장과 MPTP에 의해 세포사멸에 대한 MWE의 보호효과를 규명하고자 하였다. MWE는 MPTP의 신경독성에 의한 SH-SY5Y 세포생존율의 감소 및 세포 내 ROS 생성을 유의하게 억제시켰다. 또한 MWE는 MPTP로 유발된 SH-SY5Y 세포주의 세포자멸사(apoptosis)를 세포사멸 실행 단백질인 caspase-3 활성을 억제하여 완화시켰다. 이상의 연구결과들을 종합하면, MWE는 항산화 능력을 가지고 있어 ROS의 생성을 억제하고, 세포사멸과 관련 세포 내 효소계를 억제함으로써 도파민 세포의 사멸을 억제하여 파킨슨병에 유효하게 활용할 수 있을 것으로 생각된다.
결론
-
이해상충
이 연구에 영향을 미칠 수 있는 기관이나 이해당사자로부터 재정적, 인적 지원을 포함한 일체의 지원을 받은 바 없으며, 연구윤리와 관련된 제반 이해상충이 없음을 선언한다. 이 논문은 대한임상독성학회지와 다른 학회지에 동시 투고되지 않았으며 이전에 다른 학회지에 게재된 적이 없다.
Notes



- 1. Coleman C, Martin I. Unraveling Parkinson’s disease neurodegeneration: does aging hold the clues? J Parkinsons Dis 2022;12:2321-38. https://doi.org/10.3233/JPD-223363ArticlePubMedPMC
- 2. Marino BL, de Souza LR, Sousa KP, Ferreira JV, Padilha EC, da Silva CH, et al. Parkinson’s disease: a review from pathophysiology to treatment. Mini Rev Med Chem 2020;20:754-67. https://doi.org/10.2174/1389557519666191104110908ArticlePubMed
- 3. Mustapha M, Mat Taib CN. MPTP-induced mouse model of Parkinson’s disease: a promising direction of therapeutic strategies. Bosn J Basic Med Sci 2021;21:422-33. https://doi.org/10.17305/bjbms.2020.5181ArticlePubMedPMC
- 4. Gnanasegaran N, Govindasamy V, Mani V, Abu Kasim NH. Neuroimmunomodulatory properties of DPSCs in an in vitro model of Parkinson’s disease. IUBMB Life 2017;69:689-99. https://doi.org/10.1002/iub.1655ArticlePubMed
- 5. Ko JH, Lee JH, Choi B, Park JY, Kwon YW, Jeon S, et al. Neuroprotective effects of Gagam-Sipjeondaebo-Tang, a novel herbal formula, against MPTP-induced Parkinsonian mice and MPP+-induced cell death in SH-SY5Y cells. Evid Based Complement Alternat Med 2018;2018:2420809. https://doi.org/10.1155/2018/2420809ArticlePubMedPMC
- 6. Kim TK, Yong HI, Lee JH, Cha JY, Kang MC, Jung S, et al. Development of new technology for functional materials for edible insects as alternative food. Korean Soc Food Sci Anim Resour 2021;10:31-43.
- 7. Nam JH, Kim D, Hyun JY, Jin HJ, Choi YS, Cho JH, et al. Current status and future prospects of the insect industry as an alternative protein source for animal feed. J Korean Soc Food Sci Nutr 2022;51:395-402. https://doi.org/10.3746/jkfn.2022.51.5.395Article
- 8. Lee JH, Moon BC, Nam HH, Kim JS. Analysis of trends in patents on insect-derived medicinal materials for skin diseases. Korea J Herbol 2020;35:39-46. https://doi.org/10.6116/kjh.2020.35.2.39Article
- 9. Lee JH, Moon BC, Kim JS. Analysis of trends in patents on insect medicinal materials for brain diseases. Korean Herb Med Inf 2020;8:1-12. https://doi.org/10.22674/KHMI-8-1-1Article
- 10. Kim SH, Shon JY, Park JS, Kim JW, Kang JH, Yun EY, et al. Change in dietary intake and nutritional status using mealworms as hospital meal in postoperative patients. J Korean Diet Assoc 2016;22:292-309. https://doi.org/10.14373/JKDA.2016.22.4.292Article
- 11. Yoon OY, Hwang JS, Goo TW, Kim MA, Chung MY. Composition for prevention or treatment of obesity comprising Tenebrio molitor larva extract or Tenebrio molitor larva suspension (KR patent 10-2016-0041138) [Internet]. Daejeon: Korean Intellectual Property Office; 2016 [cited 2023 Dec 5]. Available from: https://doi.org/10.8080/1020140134331Article
- 12. Baek M, Seo M, Kim M, Yun EY, Hwang JS. The antioxidant activities and hair-growth promotion effects of Tenebrio molitor larvae extracts (TMEs). J Life Sci 2017;27:1269-75. https://doi.org/10.5352/JLS.2017.27.11.1269Article
- 13. Yu JM, Jang JY, Kim HJ, Cho YH, Kim DI, Kwon OJ, et al. Antioxidant capacity and Raw 264.7 macrophage anti-inflammatory effect of the Tenebrio molitor. Korean J Food Preserv 2016;23:890-8. https://doi.org/10.11002/kjfp.2016.23.6.890Article
- 14. Kim JH, Kim EY, Chung KJ, Lee JH, Choi HJ, Chung TW, et al. Mealworm oil (MWO) enhances wound healing potential through the activation of fibroblast and endothelial cells. Molecules 2021;26:779. https://doi.org/10.3390/molecules26040779ArticlePubMedPMC
- 15. Yi HY, Chowdhury M, Huang YD, Yu XQ. Insect antimicrobial peptides and their applications. Appl Microbiol Biotechnol 2014;98:5807-22. https://doi.org/10.1007/s00253-014-5792-6ArticlePubMedPMC
- 16. Nam ST, Kim DH, Lee MB, Nam HJ, Kang JK, Park MJ, et al. Insect peptide CopA3-induced protein degradation of p27Kip1 stimulates proliferation and protects neuronal cells from apoptosis. Biochem Biophys Res Commun 2013;437:35-40. https://doi.org/10.1016/j.bbrc.2013.06.031ArticlePubMed
- 17. Kim DH, Lee IH, Nam ST, Hong J, Zhang P, Hwang JS, et al. Neurotropic and neuroprotective activities of the earthworm peptide Lumbricusin. Biochem Biophys Res Commun 2014;448:292-7. https://doi.org/10.1016/j.bbrc.2014.04.105ArticlePubMed
- 18. Seo M, Goo TW, Chung MY, Baek M, Hwang JS, Kim MA, et al. Tenebrio molitor larvae inhibit adipogenesis through AMPK and MAPKs signaling in 3T3-L1 adipocytes and obesity in high-fat diet-induced obese mice. Int J Mol Sci 2017;18:518. https://doi.org/10.3390/ijms18030518ArticlePubMedPMC
- 19. Navarro Del Hierro J, Cantero-Bahillo E, Fernandez-Felipe MT, Garcia-Risco MR, Fornari T, Rada P, et al. Effects of a mealworm (Tenebrio molitor) extract on metabolic syndrome-related pathologies: in vitro insulin sensitivity, inflammatory response, hypolipidemic activity and oxidative stress. Insects 2022;13:896. https://doi.org/10.3390/insects13100896ArticlePubMedPMC
- 20. Di Mattia C, Battista N, Sacchetti G, Serafini M. Antioxidant Activities in vitro of Water and Liposoluble Extracts Obtained by Different Species of Edible Insects and Invertebrates. Front Nutr 2019;6:106. https://doi.org/10.3390/insects13100896ArticlePubMedPMC
- 21. Yu MH, Lee HS, Cho HR, Lee SO. Enzymatic preparation and antioxidant activities of protein hydrolysates from Tenebrio molitor larvae (Mealworm). J Korean Soc Food Sci Nutr 2017;46:435-41. https://doi.org/10.3746/jkfn.2017.46.4.435Article
- 22. Tang Y, Debnath T, Choi EJ, Kim YW, Ryu JP, Jang S, et al. Changes in the amino acid profiles and free radical scavenging activities of Tenebrio molitor larvae following enzymatic hydrolysis. PLoS One 2018;13:e0196218. https://doi.org/10.1371/journal.pone.0196218ArticlePubMedPMC
- 23. Kim JJ, Kim KS, Yu BJ. Optimization of antioxidant and skin-whitening compounds extraction condition from Tenebrio molitor larvae (mealworm). Molecules 2018;23:2340. https://doi.org/10.3390/molecules23092340ArticlePubMedPMC
- 24. Manniello MD, Moretta A, Salvia R, Scieuzo C, Lucchetti D, Vogel H, et al. Insect antimicrobial peptides: potential weapons to counteract the antibiotic resistance. Cell Mol Life Sci 2021;78:4259-82. https://doi.org/10.1007/s00018-021-03784-zArticlePubMedPMC
- 25. Hwang D, Lee SH, Goo TW, Yun EY. Potential of antimicrobial peptide-overexpressed Tenebrio molitor larvae extract as a natural preservative for Korean traditional sauces. Insects 2022;13:381. https://doi.org/10.3390/insects13040381ArticlePubMedPMC
- 26. Thiruvengadam M, Venkidasamy B, Subramanian U, Samynathan R, Ali Shariati M, Rebezov M, et al. Bioactive compounds in oxidative stress-mediated diseases: targeting the NRF2/ARE signaling pathway and epigenetic regulation. Antioxidants (Basel) 2021;10:1859. https://doi.org/10.3390/antiox10121859ArticlePubMedPMC
- 27. Abdul-Latif R, Stupans I, Allahham A, Adhikari B, Thrimawithana T. Natural antioxidants in the management of Parkinson’s disease: review of evidence from cell line and animal models. J Integr Med 2021;19:300-10. https://doi.org/10.1016/j.joim.2021.03.007ArticlePubMed
- 28. Zielinska E, Baraniak B, Karas M. Antioxidant and anti-inflammatory activities of hydrolysates and peptide fractions obtained by enzymatic hydrolysis of selected heat-treated edible insects. Nutrients 2017;9:970. https://doi.org/10.3390/nu9090970ArticlePubMedPMC
- 29. Gupta A, Kumar A, Kulkarni SK. Targeting oxidative stress, mitochondrial dysfunction and neuroinflammatory signaling by selective cyclooxygenase (COX)-2 inhibitors mitigates MPTP-induced neurotoxicity in mice. Prog Neuropsychopharmacol Biol Psychiatry 2011;35:974-81. https://doi.org/10.1016/j.pnpbp.2011.01.017ArticlePubMed
- 30. Niranjan R, Mishra KP, Thakur AK. Inhibition of cyclooxygenase-2 (COX-2) initiates autophagy and potentiates MPTP-induced autophagic cell death of human neuroblastoma cells, SH-SY5Y: an inside in the pathology of Parkinson’s disease. Mol Neurobiol 2018;55:8038-50. https://doi.org/10.1007/s12035-018-0950-yArticlePubMed
 PubReader
PubReader ePub Link
ePub Link Cite
Cite